
Oligodendrositlerin ve Schwann hücrelerinin ana işlevi miyelin oluşumudur. Miyelin, aksonal segmentler için bir yalıtkan görevi görür ve 200 m/s’ye kadar yüksek nöronal iletim hızı için bir ön koşuldur. Glial hücrelerin aksonlara bağlanması omurgasızlarda da bulunmuştur. Omurgalı Rimac hücrelerine benzer şekilde yutan aksonal hücreler, çoğu omurgasızda bulunur. Oligodendrositler ve Schwann hücreleri tarafından miyelin oluşumu, yaklaşık 400 milyon yıl önce omurgalıların evrimsel bir icadıdır. Çenesiz balıklar hariç tüm omurgalılar (jambon ve taş balığı) oligodendrositler içerir. Miyelinin gelişimi, omurgalıların, özellikle de sinir sistemlerinin evrimini hızlandırdı. Çoğu sinirbilimci bile, oligodendrositlerin omurgalı gelişimindeki önemini takdir etmez. Beynin gelişmesiyle birlikte insanlarda nöron sayısının 100 milyara çıktığı biliniyor gibi görünse de, tüm bu nöronların sadece miyelin sayesinde girift bir şekilde birbirine bağlanabileceği net değildir. Bu, aşağıdaki örnekle kolayca gösterilebilir. Sinir iletim hızını arttırmanın bir stratejisi miyelin oluşumu, diğeri ise aksonun çapını arttırmaktır.
Kalamarın içindeki dev aksonların çapı 1 mm’dir ve miyelinli motor aksonlara benzer iletim hızlarına ulaşırlar. İnsan optik siniri, yüksek hızda hareket eden yaklaşık bir milyon miyelinli akson içerir. Mürekkep balıklarının 1 mm çapındaki dev akson salınımı, 0,75 μm’lik bir akson çapına eşittir.
Oligodendrositlerin morfolojisi
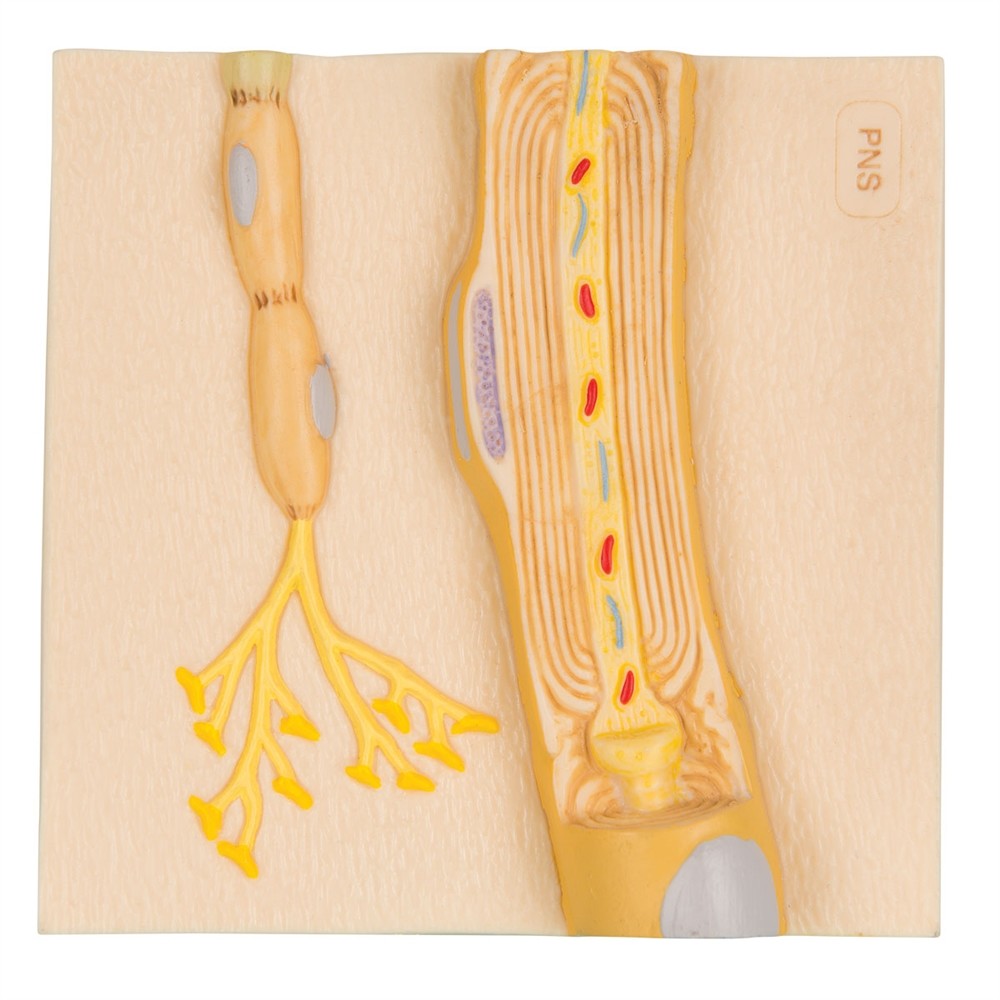
Tüm beyaz madde izleri, miyelin oluşturan oligodendrositler içerir. Oligodendrositler de gri maddede bulunur. Oligodendrositler, merkezi sinir sisteminde miyelin oluşturan hücreler olarak bilinirken, miyelin kılıfı ile doğrudan ilişkili olmayan oligodendrositler de vardır. Uydu oligodendrositler, tercihen gri maddede bulunur ve astrositlere benzer şekilde iyonik homeostazı düzenlemek için şimdiye kadar bilinmeyen işlevlere sahiptir. Sadece sıçan, fare ve insan retinalarında miyeloid oligodendrosit bulunmazken, tavşan ve civciv retinalarında kısmen miyelin bulunur. Miyelin oluşturan oligodendrositler, miyelin kısmıyla ilişkili birkaç işleme (40’a kadar) sahiptir. Bu bölümlerin her biri birkaç yüz mikrometre uzunluğundadır ve endosimbiyont olarak da adlandırılır. Segmentler, Ranvier düğümleri olarak bilinen ve 1 mikrondan daha az yayılan yapılar tarafından kesilir. Ganglionda akson, iç bölgelere göre miyelin ile çevrili değildir. Multimodal segmentin sonu daha fazla sitoplazma içerir ve aksonla segregasyon benzeri bağlantılar oluşturan paranodal döngüyü oluşturur. Ek olarak, astrosit süreçleri ganglionik bölgede akson zarı ile temasa geçer.
Astrositler gibi, oligodendrositler de konneksinler tarafından oluşturulan boşluklarla birbirine bağlanır. Astrositlere kıyasla oligodendrositler için belirgin connexin proteinleri vardır. Connexin proteinlerindeki mutasyonlar, miyelodistrofiye ve lökodistrofi gibi insan hastalıklarına yol açar.
oligodendrositlerin gelişimi
Kemirgenlerde miyelin oluşumu doğumla başlar ve doğumdan iki ay sonra tamamlanır. İnsanlarda fetal yaşamın ikinci yarısında başlar ve omurilikte gelişir. Doruk aktivite, doğum sonrası ilk yıldan 20 yaşına kadar sürer. Genellikle daha büyük aksonların daha kalın miyelin oluşturduğu bildirilmektedir. Gelişme sırasında, oligodendrositler, serebrumun lateral ventriküllerinin subventriküler bölgesi veya serebellumun dördüncü ventrikülü gibi subventriküler bölgede bulunan öncülerden ortaya çıkar. Omurilikte oligodendrositler, nöral tüpün ventral bölgelerinden kaynaklanır ve optik sinirin üçüncü ventrikülünden sinire göç eder. Hedeflerine göç eden ve daha olgun oligodendrositlere farklılaşan oligodendrosit progenitör hücrelerdir. Oligodendrosit progenitör hücrelerinin çoğalması çoğunlukla nöronlardan değil, astrositlerden salınan trombosit türevli büyüme faktörü (PDGF) veya fibroblast büyüme faktörü (FGF) gibi bir dizi büyüme faktörü tarafından kontrol edilir. Üstelik dahili saat, hücre bölünmesini hesaplamakla kalmaz, aynı zamanda zamanı da algılar. Bu nedenle, endojen ve çevresel mekanizmalar, miyelogenez için gerekli olan uygun miktarda oligodendrositleri kontrol eder. Aşırı üretilen oligodendrositler (normal koşullar altında meydana gelir) apoptoz ile elimine edilir. Oligodendrosit progenitör hücrelerinin çoğalması çoğunlukla nöronlardan değil, astrositlerden salınan trombosit türevli büyüme faktörü (PDGF) veya fibroblast büyüme faktörü (FGF) gibi bir dizi büyüme faktörü tarafından kontrol edilir. Üstelik dahili saat, hücre bölünmesini hesaplamakla kalmaz, aynı zamanda zamanı da algılar. Bu nedenle, endojen ve çevresel mekanizmalar, miyelogenez için gerekli olan uygun miktarda oligodendrositleri kontrol eder. Aşırı üretilen oligodendrositler (normal koşullar altında meydana gelir) apoptoz ile elimine edilir. Oligodendrosit progenitör hücrelerinin çoğalması çoğunlukla nöronlardan değil, astrositlerden salınan trombosit türevli büyüme faktörü (PDGF) veya fibroblast büyüme faktörü (FGF) gibi bir dizi büyüme faktörü tarafından kontrol edilir. Üstelik dahili saat, hücre bölünmesini hesaplamakla kalmaz, aynı zamanda zamanı da algılar. Bu nedenle, endojen ve çevresel mekanizmalar, miyelogenez için gerekli olan uygun miktarda oligodendrositleri kontrol eder. Aşırı üretilen oligodendrositler (normal koşullar altında meydana gelir) apoptoz ile elimine edilir.
Hâlâ astrositlere ve oligodendrositlere yol açabilen oligodendrosit progenitör hücreler, yalnızca gelişim sırasında mevcut olmakla kalmaz, aynı zamanda yetişkin oligodendrosit progenitör hücreler olarak adlandırılan olgun beyinde de bulunur. Multipl skleroz gibi demiyelinizan hastalıklarda remiyelinizasyon kaynağıdır. Bu birincil hücrelerin tanımlanmasına yardımcı olan, transkripsiyon faktörü Olig-2 veya NG2 proteoglikan gibi birkaç farklı belirteç vardır. Bu NG2-pozitif hücreler son zamanlarda çok ilgi gördü. Astrositlere ve oligodendrositlere dönüşme potansiyeline sahip olmalarına rağmen, ana yolun oligodendrosit soylarıyla sınırlı olduğu görülmektedir. Bu yetişkin progenitör hücreler, aksonlarla etkileşime giriyor gibi görünmektedir. Glutamat reseptörlerini eksprese ederler ve aktiviteye bağlı bir şekilde glutamat salan aksonların aktivitesini algılarlar. Bu, aksonların oligodendrosit progenitör hücrelerinin farklılaşmasını nasıl kontrol ettiğine dair potansiyel bir mekanizma gibi görünmektedir.
Schwann hücreleri
Schwann hücreleri, periferik sinir sistemindeki oligodendrositlerin hücresel karşılığıdır. Oligodendrositlere benzer bir miyelin kılıfı oluştururlar. Oligodendrositlerin aksine, her bir Schwann hücresi yalnızca bir aksonal segment ile ilişkilidir. Oligodendrositlerin ve Schwann hücrelerinin miyelinleri benzer bir yapıya sahipken aynı protein grubundan oluşmazlar. Merkezi ve periferik miyelin, temel bir miyelin proteinini paylaşırken, periferik sinir sistemi miyelinle ilişkili bir glikoprotein veya lipoproteinden yoksundur, ancak P0 ve PMP22 proteinlerini ifade eder. Gelişim sırasında Schwann hücreleri, farklılaşmamış göçmen nöral krest hücrelerinden türetilir. Olgunlaşmamış Schwann hücreleri, miyelinleşmeyen Schwann hücreleri üretir. İkincisi, miyelin oluşturmadan birkaç aksonu gevşek bir şekilde yutar.
Sempatik ve parasempatik duyu ganglionlarındaki sinirlerin hücre gövdeleri, uydu hücreleri olarak bilinen düz, kılıf benzeri hücrelerle çevrilidir. Nöromüsküler kavşaktaki akson terminalleri ayrıca periferal glia gibi özelleşmiş glial hücrelerle kaplıdır.
İçindekiler
miyelin kılıfları
Miyelin kılıfı, aksonun oligodendrositler veya Schwann hücrelerinin işlemleriyle sarılmasıyla oluşturulur. Hücre içi alan çok kompakttır, yalnızca 30 angstromu kapsar ve elektron mikroskobunda ana yoğun çizgi olarak adlandırılan tek bir çizgi olarak görünür. Lipit çift tabakasının dış yüzeyi, hücre dışı boşlukla ayrılmış ayrı bir çizgi olarak gösterilir. Bu nedenle, bu çift dönem çizgisi olarak tanımlanır. Bu yoğun stres nedeniyle miyelin tamamen hidratlanır ve kuru kütlesi yaklaşık %70 yağ ve %30 protein içerir. Sadece miyelinde bulunan ve bu yapının oluşumu için gerekli olan birkaç spesifik protein vardır. Miyelin merkezi sinir sisteminin ana proteinleri, miyelinle ilişkili glikoprotein (MAG), miyelin temel proteini (MBP), miyelin oligodendrosit glikoproteini (MOG), lipoprotein (PLP)/DM20 ve PMP22’dir. Bu proteinler yalnızca miyelin oluşturan hücreler, yani merkezi sinir sistemindeki oligodendrositler veya periferik sinir sistemindeki Schwann hücreleri tarafından üretilir ve bu nedenle miyelin hücreleri için mükemmel belirteçler olarak hizmet eder. Miyelin katmanları içinde, Schmidt-Lintermann fissürleri adı verilen visseral bir vakuol içeren bir tür yol vardır. Bunlar miyelin için beslenme desteği sağlar.
Tüm omurgalı aksonları tamamen miyelinli değildir, ancak genel olarak 1 μm’den büyük aksonlar miyelinle doludur. Son çalışmalar, aksonların miyelin kılıfının kalınlığını belirleyen oligodendrositlere bir sinyal sağladığını göstermektedir. Akson tarafından sağlanan önemli bir sinyal mekanizması, oligodendrositler tarafından eksprese edilen ErbB reseptörü tirozin kinazlara bağlanan büyüme faktörü neuregulin-1’dir. Schwann hücreleri benzer bir sinyal mekanizmasına sahiptir. Bu etkileşim, miyelin kılıfın yanı sıra akson çapı ile akson çapı arasında belirli bir oran ile sonuçlanır; Bu oran genellikle 0,6 ile 0,7 arasındadır.
Miyelin hücrelerinin aksonlar için metabolik destek sağladığı uzun zamandır öne sürülmüştür. Piruvat veya laktat gibi gliadan türetilen glikolitik ürünlerin salınması ve akson tarafından alınması beklenebilir. Bu, periferik sinir sistemi için daha önemli olabilir çünkü somadaki metabolitler, büyük hayvanlarda 1 metreden daha uzun mesafeler kat etmek zorundadır.
Miyelin sinir iletimini sağlar
Ranvier düğümü, tuz iletimi kelimesinin (“atlamak” anlamına gelen Latince “saltari” kelimesinden), yani yalnızca düğümde aksiyon potansiyelleri üretmesine izin veren yüksek yoğunluklu bir sodyum kanalı içerir. Böylece, aksiyon potansiyelleri sadece bir düğümde tetiklenir, sonra pasif olarak yayılır ve böylece bir sonraki aksiyon potansiyelinin üretildiği bir sonraki düğüme hızla hareket eder. Böylece aksiyon potansiyeli bir düğümden diğerine iletilir. Bu sadece daha hızlı olmakla kalmaz, aynı zamanda daha az enerji gerektirir çünkü sodyum iyonları sadece ganglionda birikir ve Na+/K+ aktivitesi nedeniyle hücre dışı boşluğa geri taşınması gerekir. Miyelin oluşumundan önce, sodyum kanalları akson boyunca rastgele dağılır. Bununla birlikte, glial oklüzyon sırasında, sodyum kanalları alanda daha sonra çalışan düğümler haline gelen gevşek yığınlar oluşturmaya başlar. Daha sonra, sıkıştırılmış miyelin oluşumundan sonra, sodyum kanalları miyelin kılıfının altındaki zardan kaybolur ve sadece ganglionda toplanır. Bu düzenek, miyeloid hücre zarı ile glonmedin, nörofazin ve NCAM gibi hücre adezyon moleküllerini içeren aksonal zar arasındaki protein etkileşimleri ile desteklenir. K+ kanalları düğüm bölgesinde daha az yoğunlaşmıştır.
Miyelin hücreleri ve hastalıkları
Multipl skleroz, oligodendrositleri etkileyen en yaygın hastalıktır. Beynin ve omuriliğin belirli bölgelerinde miyelin kaybına bağlı olarak zayıf aksonal iletime neden olur. Remiyelinizasyon nedeniyle iyileşme meydana gelebilir, ancak sıklıkla kalıcı nörodejenerasyonla sonuçlanan nüksler meydana gelir. Oligodendrosit kaybının ana nedeni henüz bilinmemektedir. Demiyelinize alan açıkça infiltre edici lenfositler, makrofajlar gibi enflamatuar hücreler ve aktive edilmiş mikroglia içerir. Bu hücreler hasar zincirini güçlendirebilir ve hatta başlatabilir. Merkezi sinir sisteminin diğer kalıtsal demiyelinizan hastalıkları Pelizaeus-Merzbacher hastalığı, Pelizaeus-Merzbacher benzeri hastalık ve diğer lökodistrofi biçimleridir. Genetik olarak belirlenmiş hastalıkların çoğu, miyelin proteinleri veya konneksinlerdeki mutasyonlarla ilişkilidir ve moleküler varlıklar boşluk bağlantılarını oluşturur. Miyelin, Schwann hücresi veya boşluk bağlantı proteinlerindeki benzer CNS veya merkezi sinir sistemi mutasyonları, Charcot-Marie diş hastalığı gibi nöropatilere yol açar. Bu, periferik miyelin oluşumunun da omurgalıların hayatta kalması için gerekli olduğunu göstermektedir.
kaynak:
Britanya
yazar: Tuncay Bayraktar
Diğer gönderilerimize göz at
[wpcin-random-posts]