
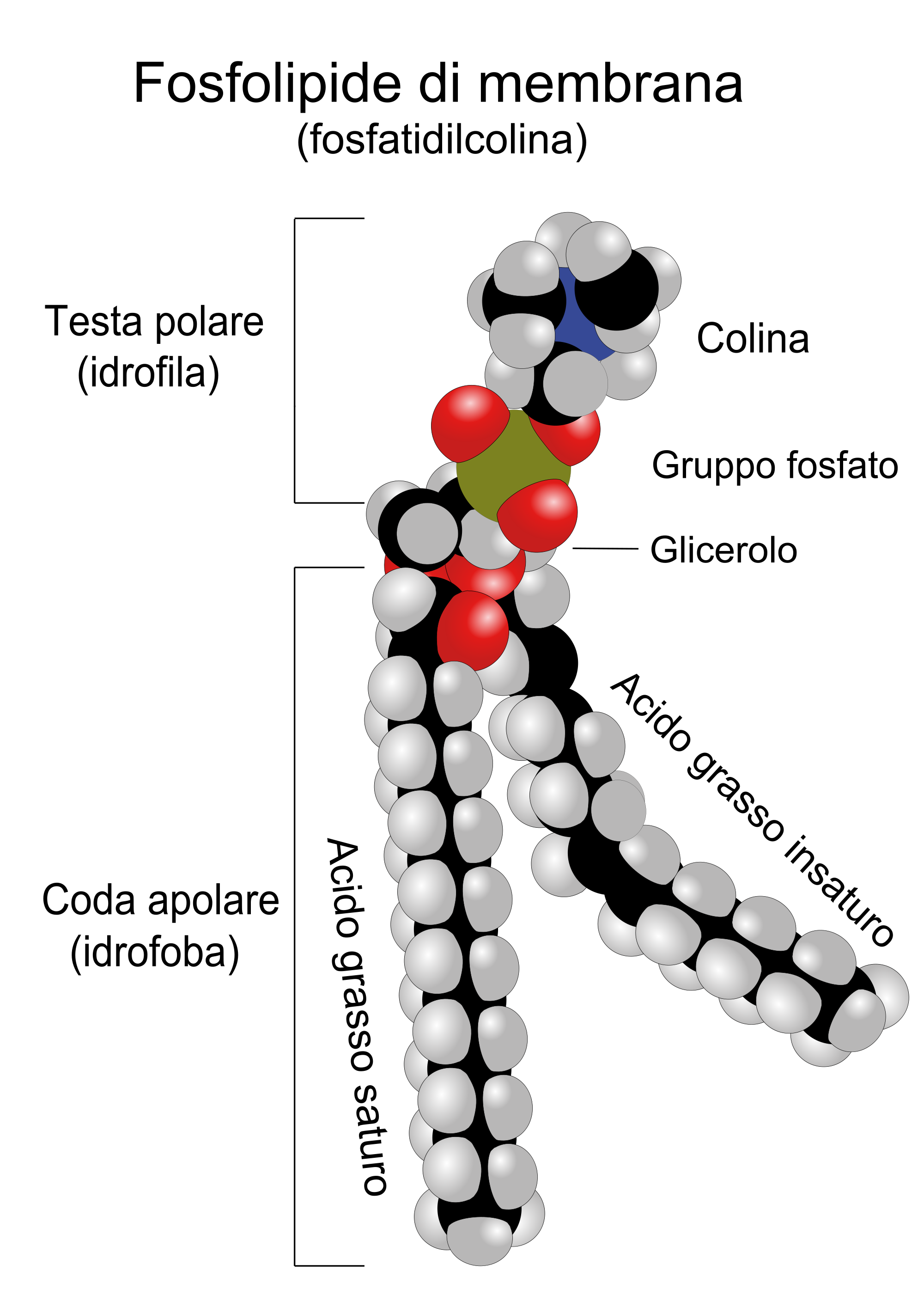
Araştırmacılar onlarca yıldır hücre zarının dikkate değer fonksiyonlarını ve yapısını incelediler. Membran işlevlerini çözmek, zarın yapısını anlamaktan ve ardından çeşitli özelliklerini anlamaktan oluşur. Örneğin, geçirgenlik çalışmaları sıvı ve yağda çözünen maddelerin hücreler arasında ve hücre dışı ortamlara diğer maddelere göre daha kolay hareket edebildiğini göstermiştir. Geçmiş yıllarda araştırmacılar, hücre zarının lipid içermesi gerektiği ve ancak bu şekilde yağda çözünen maddelerin zarda çözünerek hücre içine geçebileceği sonucuna varmışlardır. Mevcut bilgilerimize yönelik bu önemli ilk adım; 1930’ların sonlarında Princeton Üniversitesi’nden JF Danielli ve Londra Üniversitesi’nden H. Davson tarafından okuldan atıldı. Danieli ve Daphson, hidrofobik zarın iki fosfolipit tabakasından oluştuğu ve fosfolipidin polar (hidrofilik) kısmının zarın dış tarafında ve polar olmayan (hidrofobik) kısmın iç kısımda yer aldığı fikrini formüle ettiler. zarın yanında bulunur ve iki kısımdan oluşur. Hidrofilik ve hidrofilik ilişkisindeki yapı oldukça dayanıklı ve esnek olacaktır. Aslında, şimdi “lipozomlar” olarak bilinen kürecikler, fosfolipid yapılardan oluşur. Bu fosfolipid bazlı globüller, su ile karıştırılsa bile bir süre sonra kendiliğinden oluşur. Hücre zarının mikroskobik görüntüleri, lipozomların mikroskobik görüntülerine çok benzer ve lipitlerin, ilk organizmaların değişmesini mümkün kılan çift katmanlı boşluklar oluşturmalarına izin veren bir yapıya sahip olmaları muhtemeldir.
Fosfolipid çift tabakalı model, dayanıklılığın, esnekliğin ve yağın hücre zarından geçiş kolaylığının mekanizmasını açıklasa da, iyonların ve kimyasalların hücre zarından seçici geçişi için kesin bir açıklama sağlayamadı. Dufson ve Danieli, zarın her iki tarafının da proteinle çevrili olabileceği fikrini öne sürdüler. Yüklü proteinden yapılmış gözenekler, küçük moleküllerin ve iyonların hücre zarından geçmesine izin verir. Ancak modelin bu kısmının hatalı olduğu gösterildi.
İçindekiler
Mozaik akıcı zar modeli
1972’de California Üniversitesi’nden SJ Singer ve Salk Enstitüsü’nden GL Nicholson, neredeyse tüm dünyada kabul gören sıvı mozaik zar modeli hipotezini ortaya attılar. Bu modele göre, Daniele ve Daveson’ın fosfolipid çift tabakasına ilişkin görüşü, fosfor sferoitlerin hidrofilik başlarının zarın her iki tarafında sulu ortama baktığı, hidrofobik kuyrukların ise zarın iç kısmına baktığı görüşüyle birleşir. Ancak sıvı mozaik modelde proteinlerin dizilişi oldukça farklıdır. Proteinler, zarı kaplamak yerine, çeşitli şekillerde yaşamsal işlevlerin büyük çoğunluğunu gerçekleştiren zar tarafından yutulurlar.
Yüzeyi çevreleyen proteinlere periferik proteinler denir. İç yüzeyde olan şey, genellikle dış yüzeyde olandan önemli ölçüde farklıdır. Bu asimetri özelliği, zarın diğer asimetrik özelliklerinden sadece bir tanesidir. Örneğin fosfolipidler, çift tabakanın iç ve dış yarısında tamamen farklı bir düzenleme gösterir. Karbonhidrat grubu bağları, zarın yalnızca dış yüzeyinde bulunurken, proteinlerin çoğu, bir kısmı hemen hemen her zaman içte veya dışta olmak üzere, lipid çift tabakasına (integral proteinler) dahil edilir. Entegre proteinler birçok farklı bağlayıcı görüntüleyebilir: bazıları çift tabakanın içine tamamen gömülmüş olabilir. Bazılarının yüzeye doğru çıkıntı yapan parçaları vardır. Bazıları yağlı çekirdeğin dış yarısında, diğerleri iç yarısında bulunur. Diğerleri yağ tabakası boyunca uzanır. Beklendiği gibi, hidrofilik amino asitler (bunlar polar veya elektrik yüklü R gruplarıdır), protein moleküllerinin lipit çift tabakasından suya doğru çıkıntı yapan kısımlarında daha fazla bulunur. Bununla birlikte, kısmen hidrofobik (polar olmayan) amino asitler, lipit çift tabakasında bol miktarda bulunur. Aslında, bir zardaki proteinler içindeki hidrofobik ve hidrofobik amino asitlerin konumu, proteinin hangi kısmının zarda kalacağını ve hangi kısmının çıkacağını veya proteinin tamamlayıcı mı yoksa periferik mi olacağını belirler. Varlığı yalnızca sıvı mozaik zar modeliyle belirlenen entegre proteinler artık kriyoskopik mikroskopi ve diğer tekniklerle doğrulanıyor.
Sıvı-membran modeline göre membran yapısı statik değildir. Her lipid molekülü, zar çizgisi boyunca sola ve sağa hareket edebilir. Bu nedenle, belirli bir zamanda belirli bir konumda olan bir molekül, birkaç saniye sonra tamamen farklı bir konumda olabilir. Lipit hareketliliği, doymamış fosfolipidler açısından zengin ve kolesterol içermeyen zarlarda daha fazladır. Böyle bir zar için saniyede 2 μm hız mümkündür. Böylesine inanılmaz bir hareket, yalnızca 2 mikrometre yüksekliğindeki birçok organizmada halihazırda bulunabilir. (Ör: Escherichia coli). Kolesterol mevcutsa, kolesterol komşu fosfolipidlere zayıf bir şekilde bağlanarak iki etkiye neden olabilir. Fosfolipid zarı çoğunlukla doymuşsa, kolesterol, fosfolipitlerin çok sıkı ve düzenli bir şekilde paketlenmesini önler, bu da sertleşmeye ve katı halde kristalleşmeye yol açar. Bununla birlikte, eğer fosfolipitler çoğunlukla doymamışsa (sonuç olarak, hidrofobik kuyruktaki kıvrımlar fosfolipitleri gevşek tutar), bir kolesterol molekülü bu boşlukları doldurabilir, komşu fosfolipitlere yapışabilir ve böylece birbiriyle kaynaşabilir. Bitki hücre zarlarında kolesterol yoktur. Bunun yerine, dayanıklılık hücre duvarı tarafından sağlanır.
Kolesterol konsantrasyonu ve doyma derecesi, gerekli esneklik miktarına bağlı olarak türler arasında ve hatta aynı organizma içindeki dokular arasında büyük farklılıklar gösterir. Ancak tıpta insan beslenmesinde bol miktarda bulunan kolesterol ve doymuş yağların hücre zarının sertleşmesinde çok önemli rol oynadığı ve bu nedenle özellikle atardamarlarda sertleşmeye neden olacağı ileri sürülmektedir. duvarlar. Arterlerin sertleşmesi (damarların sertleşmesi), kalp çarpıntısının ve kalp hastalığının en önemli nedenidir. Hücre zarındaki proteinler bir dereceye kadar yanal difüzyonla hareket edebilir; Ancak bu hareketler yağdan daha azdır. Tam hareket serbestliği, zardaki proteinlerin spesifik fonksiyonel gereklilikleriyle uyumlu değildir. Örneğin, sinir uyarılarının bir hücreden diğerine iletilmesi için sinir hücresi zarındaki bazı proteinlere ihtiyaç vardır. Sadece bir sinir hücresinin diğerine bağlandığı belirli noktalarda bulunurlar; Diğer pozisyonlarda işlerini yapamayacakları yerlerde değiller. Benzer şekilde, sodyum iyonlarını bağırsağı kaplayan hücrelere pompalamaktan sorumlu proteinler, hücrelerin yalnızca bir tarafındaki zarda bulunur. Bu taraf bağırsak lümeninden en uzak kısımdır.
Kısacası, bazı transmembran proteinler, zar akışkanlığını sınırlayan zara gömülür. Bazı durumlarda, bu yırtılma, oldukça büyük yapısal ve fonksiyonel komplekslerin kolayca ilerlemesine izin veren iki veya daha fazla içsel protein arasındaki yakın ilişkilerin sonucu olabilir. Diğer durumlarda, periferik ve proteolitik proteinler birbirine zayıf bir şekilde bağlanabilir. Lipit molekülleri hareketlerinde tamamen serbest olmasa bile, içsel proteinlere doğrudan bağlı olan lipitler, proteinlerle zayıf bağlar oluşturarak onları hareketsiz kılar. Akışkan mozaik modelinde, zardaki gözenekler, bir veya bir grup protein molekülü arasındaki kanallar olarak görselleştirilir. Lipit çift tabakası içindeki kararsız proteinlerin yanal kayması, birçok zar gözeneklerinin gözlenen hareketini açıklar. Bir proteindeki amino asitlerin farklı R gruplarını ayırt eden özellikler, gözeneklere bazı seçici özellikler verir ve gözeneklere sığacak kadar büyük bazı iyonların veya moleküllerin gözeneklerden fiilen hareket etmesine izin verir.
kaynak:
Khan Akademisi
yazar: bronzlaştırıcı tonik
Diğer gönderilerimize göz at
[wpcin-random-posts]