
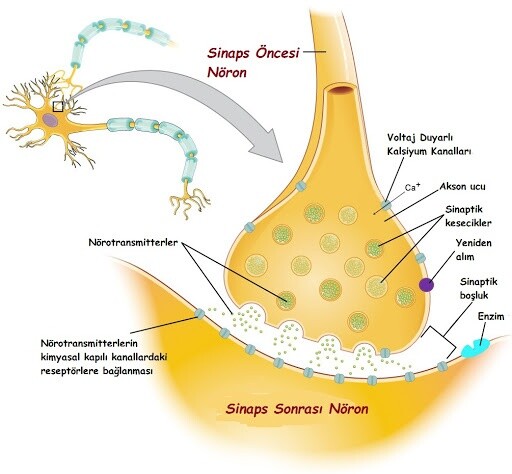
Klasik glutamaterjik sinaps, dendritik omurga ağacının tepesinde bulunur. Omurga, dendritik bir gövdeden çıkıntı yapan bir boyun ve bilginin gerçekten alındığı bir kafadan oluşur. Glutamat sinapsının genel bir tanımı, postsinaptik omurga ile yarıktan bakan bir presinaptik düğmeyi içerir. Presinaptik düğmenin postsinaptik omurganın karşısındaki bölgesi, glutamatla dolu veziküller içerir ve aktif bölge (AZ) olarak adlandırılır. 10 ila 20 vezikülden oluşan bir grup, presinaptik zara, salınmaya hazır veziküllere giren bir protein kompleksi olan SNARE kompleksi (Çözünür NSF bağlayıcı proteinler) tarafından bağlanır.
Presinaptik spike’ın gelişi vezikül füzyonunu ve Ca2+ akışından sonra SNARE kompleksi tarafından aktive edilen gözenek oluşumunu aktive eder. Temel tek bilgi parçasının iletilmesindeki ilk adım, SNARE kompleksi tarafından düzenlenen glutamat vezikül salımıdır, bunu takiben presinaptik yükselme gelir. Yüksekliği iletilen nöral bilginin çekirdek biti olarak düşünürsek, EPSP sinaptik olarak kodlanmış bilginin çekirdek biti olarak düşünülebilir.
Eklenen vezikülleri içeren presinaptik yüzey, postsinaptik yüzeyden 20 nm’lik bir mesafe (yarık) ile ayrılır. Sinaptik yarık, presinaptik vezikülden salınan glutamat moleküllerinin Brownian hareketi ile yayıldığı hacimdir. Presinaptik dikenlerin Ca2+ kompleksi ve SNARE yoluyla gelmesi, vezikül ile presinaptik zar arasında bir gözenek oluşumuna yol açar. Bu gözenek, glutamat moleküllerinin kesecikten sinaptik yarığa geçerken izledikleri yoldur.
Genel bir enine kesit varsayarsak, kortikal veya hipokampal glutamaterjik sinapsların çapı 0,2–1 μm’dir. Sıfırdan dairesel bir alan ve 20 nm’lik bir yarık varsayılarak, birkaç yazar tarafından bir bilgisayar modelleme yaklaşımı kullanarak sinaptik iletimi incelemek için kullanılan silindirik bir alan hacmi elde edilir. Tüm sinaptik “silindirler” glutamat difüzyonundan muaf değildir. AZ, tüm sinapsın yalnızca bir bölümünü kaplar (ortalama yarıçap 0.11 μm), çevresel kısım ise sinaptik öncesi ve sonrası nöronları tutan lifler tarafından işgal edilir.
Postsinaptik tarafta, iki tip glutamat reseptörü, AZ’ye kabaca benzer büyüklükte bir alanda lokalizedir ve dairesel (silindirin altında) olarak kabul edilir. Bu bölge postsinaptik yoğunluk (PSD) olarak adlandırılır ve amino-3-hidroksi-5-metil-4-izoksazolipropiyonik reseptörleri (AMPA reseptörleri) ve hassas N-metil-d-aspartat reseptörlerini (NMDA reseptörleri) içeren yerleri içerir. Glutamat moleküllerini bağlamak için (ayrıca sinaptik aktivite için gerekli olan glisine). Bu iki reseptör tipi, daha sonra daha ayrıntılı olarak tartışacağımız sinaptik bilginin iletilmesinde farklı rollere sahiptir. Her iki tip de dimerlerden oluşan tetramerlerdir ve reseptörün dimerik yapısı, postsinaptik yanıtın şekillenmesinde önemli bir rol oynar.
Görünüşe göre, bilgi aktarma süreci prensipte çok basit. Presinaptik dikenin gelişi, postsinaptik reseptörleri aktive eden ve dendritik dallardan yayılan bir depolarize edici akım (EPSC) üreten ve EPSP adı verilen postsinaptik membran potansiyelinde bir anizotropiye neden olan glutamat salınımı ile vezikül füzyonu üretir. (postsinaptik yüksekliği oluşturmak için somadan) Bununla birlikte, presinaptikten postsinaptik tarafa geçişlerin herhangi biri, tüm süreci incelemeyi ve yorumlamayı çok karmaşık hale getiren bir dizi bilgi yeniden düzenlemesinden geçer.
Esasen, farklı modülasyon sistemleri, postsinaptik yanıtın bir tür karmaşık doğrusal olmayan varyansını üretir. EPSP varyansına hem presinaptik hem de postsinaptik kontrol mekanizmaları neden olur ve bu varyansın bir kısmı doğası gereği rastgele görünmektedir. Postsinaptik yanıtın çeşitliliğini ve bunun potansiyel olarak rastgele veya deterministik doğasını değiştirmeye dahil olan bazı (hepsi değil) düzenleyici mekanizmalar vardır. Bu mekanizmalar:
EPSP değişkenliği için ek sinaptik faktörler
Sınırlı sinaptik boşluğun ötesine bakıldığında, diğer birkaç faktör EPSP oluşumunu etkileyebilir. Kısaca denklemlere göre bunu söyleyebilir. Sinaps yakınındaki zar potansiyelini etkileyebilecek herhangi bir faktör, EPSP oluşumunda rol oynayabilir. Sinaptik boşluğun dışında bulduğumuz ilk önemli yapı servikal omurgadır. Sinaps ve dendrit arasındaki iletişim yoludur ve elektrik direnci hücreye (dendrit) geçen bilgi miktarını belirler. Bu nedenle dendritlerin farklı bölgeleri ile soma arasındaki bilgi akışı için boyun direnci değeri çok önemlidir. Dikenlerin şekli değişkendir ve bu nedenle biyoelektrik özellikleri ve voltaj kapılı kanalların varlığı, sinaptik bilgiyi iletme yeteneklerini daha fazla etkileyebilir.
Bazı araştırmacılara göre boyun çapı ve direnci de tek bir sinaptik olay sırasında modüle edilir. Boyun direncinin modülasyonu sonuçta somaya iletilen EPSP’nin modülasyonuyla sonuçlanır. Bununla birlikte, boyun sadece birbirine kenetlenen bilgileri dendritlere taşımaz. Ayrıca dendritin durumu ile ilgili bilgileri PSD’ye ileterek ters yönde hareket eder. Başka bir deyişle, PSD yakınlarda bulunan diğer sinapslardan gelen bilgilerin farkındadır. Dendritik aktivite aslında dendrit ve omurga başı arasında potansiyel bir fark yaratır, bu da yönü iki yapı arasındaki potansiyel farka bağlı olan net bir akımla sonuçlanır. Dendritten gelen akım, esas olarak, toplam EPSP ve NMDA reseptörü toplanmasını güçlü bir şekilde etkileyen PSD’nin yüksek giriş empedansı ile yükseltilir. O halde omurilik, dendritlerdeki zar potansiyelinin uyarılma (depolarizasyon) düzeyine bağlı olarak sinaptik bilgi iletiminin güçlü bir modülatörüdür.
Çok sayıda erişim nedeniyle (3 × 1033 × 103 -e 3 × 1043 × 104), dendritik dallanma elektriksel olarak simetrik bir bölünme değildir. Farklı dallar arasındaki potansiyel farklılıklar, farklı girdi aktivitelerinden ve meydana gelebileceği bölgelerde artan geri yayılımdan kaynaklanabilir. Geri saçılım yayılımı, dendritlerde voltaj kapılı Na+ ve/veya Ca2+ kanallarının varlığına bağlıdır. Bu kanalların varlığı ve yoğunluğu, nöronlar arasında ve aynı nörondaki farklı dendritik bölgeler arasında değişir. Sonuç olarak, potansiyel farklılıklar, dendritlerden geçen karmaşık potansiyel dalgalar oluşturabilir ve bu dalga, tek bir sinaptik olayı etkileyerek boyun empedansı yoluyla PSD’ye ulaşabilir.
Son araştırmalar, uyarıcı sinaptik aktivite tarafından üretilen potansiyel dalgaların, yeniden kablolamadan bağımsız olarak tek bir sinaptik yanıt üzerindeki potansiyel etkisini inceledi. Aktif sinapsların sayısına ve bunların ortalama ateşleme frekansına bağlı olarak, yanıt genliği, tepe seviyesi ve zirveye ulaşma süresinin karmaşık bir doğrusal olmayan şekilde değiştiği bulunmuştur. Aktif sinapsların sayısı, bir şekilde beynin bir bölgesinden gelen girdiyi taklit eder, örneğin, bir grup aktif nöron aynı nöronda aşağı yukarı eşzamanlı bir şekilde (bir uyarana yanıt olarak) ateşlenir. (dendritik alan)
Yukarıda belirtilen durum için, örneğin, hipokampal alt alandaki nöronlar, dentat girusun geniş bir alanından (birçok nöron) ve ayrıca dendritik dalların ayrı bölgelerindeki entorhinal korteks bölgelerinden eşzamanlı girdiler alabilir. Bu iki bölgenin herhangi birinden gelen nöronlar, senkronizasyon derecesine bağlı olarak ortalama bir frekans ve standart sapma ile ateşlenir. Böyle bir durum, dendritik alanda, aynı zaman periyodunda aktif olan herhangi bir sinapsı doğrudan etkileyen uyarımla ilgilenen dalgalar üretir.
Alıcı nöronun zar potansiyeli, potansiyel bir “bant” oluşturmak için iki seviye arasında salınır. Bu voltaj “aralığının” genişliği, aktif sinapsların sayısına ve bunların ortalama ateşleme frekansına bağlıdır. Belirli bir sinapsın EPSP’si, ‘bant’ içindeki herhangi bir seviyede meydana gelebilir. Denklemlere göre, EPSP’nin belirli bir sinaps için başladığı seviyeye bağlı olarak, özellikleri (genlik, tepe seviyesi, NMDA katkısı vb.) değişecektir. Bu aralıkta, VmVmEPSP’nin indükleyebileceği maksimum olasılık seviyesi olarak kabul edilebilecek bir ortalama değer ayarlamak mümkündür.
Bu ara seviye, aktif sinapsların sayısını veya ateşleme frekanslarını artırarak arttırılır (artan polarizasyon).Bu voltaj ‘aralığının’ varlığı, olası seviyelerde büyük gamalar sağlar. VmVmEPSP’nin meydana gelebileceği zamana (bant içi salınım aşaması) ve sonuç olarak bağlı olarak bireysel EPSP’nin çok güçlü bir düzenleyicisini temsil eder. Başka bir deyişle, EPSP’nin belirli bir seviye ile örtüşmesi, sinapsın ilettiği bilginin türünü ve miktarını belirler. Aktiviteleri transmembran potansiyeline bağlı olan NMDA reseptörleri, bu tip düzenlemeye özellikle duyarlıdır ve gerçekten de EPSP’nin diğer sinapsların aktivitesi ile ‘çatışma’sı, LTP ve hafıza gibi fenomenler için çok önemlidir.
(NMDA’ya dayalı) Bunlar, nöronların, çoğunlukla LTP’nin, bellek fenomenlerinin ve dendrit ağacına aktif sinaptik girdilerin üretilmesinde kabuk dedektörleri olarak hareket etmesinin altında yatan mekanizmalardır.
kaynak:
https://www.cell.com/fulltext/S0896-6273(03)00608-1
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4743082/
yazar: Özlem Güvenç Ağaoğlu
Diğer gönderilerimize göz at
[wpcin-random-posts]