
Yakın zamana kadar bilim adamlarının çoğu, karmaşık davranışların (farklı kasların sırayla koordineli hareketinden kaynaklanan çeşitli duyuları ve davranışları bütünleştiren davranışlar) refleks yaylarının etkileşimi yoluyla aracılık edildiğine inanıyordu. Her refleks yayının tek bir duyu alması ve tek bir kası (veya el çekme refleksinde olduğu gibi bir çift karşıt kası) kontrol etmesi gerekiyordu. Öğrenilmiş davranışın, örneğin bir labirentte yolunu bulan fareler gibi, bu refleks devrelerinin bir dizisinin sonucu olduğu düşünülüyordu. Bu kabul edilen davranışsal organizasyon modeli, bu yüzyılın büyük bir bölümünde akademisyenlere egemen olmuştur.
İçindekiler
Böcek uçuşu
Karmaşık davranışın zincirleme reaksiyonlardan oluştuğu fikri, 1960’larda Stanford Üniversitesi’nden Donald Wilson’ın araştırmasıyla çürütüldü. Wilson bir çekirgeyi uçuran devreler üzerinde çalışıyordu. Bu, dönüşlü dizeler olarak kabul edilmek için iyi bir adaydı. Bir grup uçan kas kanatları kaldırır. Bir süre sonra diğer gruptaki esneme reseptörleri uyarılacak ve kanatları indirecek kasılmalar başlayacaktır. Kanatlar alçalırken, kaldırma kas grubundaki gerilme reseptörleri ekstansiyondaki artışla uyarılacak ve bir süre sonra “kaldırma refleksi” devreye girecektir. Ardından kanatlar kaldırılacak ve bu sefer inen kas grubundaki alıcılar bir “aşağı doğru refleks” başlatmak üzere uyarılacak ve bu sonsuza kadar devam edecek.
Devreyi izleyerek, Wilson gerekli gerilme reseptörlerini keşfetti ve reseptörlerden pektoral ganglionlara giden sinirleri kesti. Wilson, başlatıcı elemanları devreden çıkararak davranışın ortadan kalkacağını umuyordu. Ancak çekirge, neredeyse normal uçuşuna devam ederek onu şaşırttı. Ancak operasyonun bir etkisi oldu.
Kanat çırpma hızı biraz düşürüldü ve koordinasyon bir miktar geriledi. Ancak davranış devam etti. Sonraki araştırmalar, çekirgenin esneme reseptörleri olmadan bile uçabilme yeteneğinden başka bir elektrik devresinin sorumlu olduğunu ortaya çıkardı. Pektoral gangliyonlarda yer alan bir motor program ve gerinme reseptörlerinden herhangi bir dış uyarana ihtiyaç duymadan uçmak için gerekli komutları üreten bir devreydi. Tabii ki, bu doğal olarak ayarlandı.
Wilson’ın keşfinden bu yana omurgalılarda ve omurgasızlarda incelenen karmaşık, ritmik veya sıralı davranışların çoğu, bazı özel bağımsız çevreler tarafından bulundu. Bu devreler kas hareketlerini doğru sırada ve zamanlamada düzenler. Refleks zincirlerine dayanan hiçbir karmaşık davranış bilinmemektedir. Motor programların kontrol ettiği davranışların listesi beslenmeyi, yürümeyi, yüzmeyi, tırmalamayı, seslendirmeyi, yuva yapmayı, saldırmayı, kur yapmayı ve çiftleşmeyi içerir.
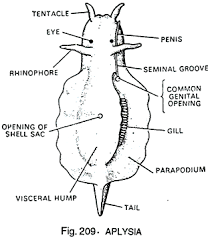
Aplicia’da Beslenme
Araştırma tekniklerinin gelişmesiyle birlikte ilgi, hücrelerin karmaşık davranışlardan sorumlu devrelerle nörofizyolojik haritalanmasına odaklandı. Günümüzde en iyi anlaşılan motor program ağları, yer değiştirme hareketinde gördüğümüz gibi, ritmik ve tekrarlayan hareketleri içerir. Pratik gereklilik nedeniyle araştırmalar, tam olarak dakikalar hatta saatler süren davranış türlerine odaklanmıştır. Bu şekilde, bilim adamlarının aynı devre üzerindeki iki hücreyi konumlandırmak, aralarındaki iletişimin doğasını belirlemek, aynı anda aktivitelerini kaydetmek ve son olarak (genellikle boya enjekte ederek) karakterize etmek için yeterli zamanı olabilir. Doğal olarak, bir devrede ne kadar çok hücre varsa ve aralarındaki ilişki ne kadar ayrıntılıysa, devreyi haritalandırmak ve anlamak o kadar fazla zaman alır.
Aplysia’daki beslenme kontrol devresi, neredeyse tamamen haritalanmış bir davranış yoluna iyi bir örnektir. Bu devre, nispeten karmaşık davranış bağlamında bilgi akışındaki dört adımı (tanıma, iletme, işleme ve yanıt) açıklar. Alglerle beslenen Aplysia’nın kafasında potansiyel yiyecekleri tespit eden kemo-duyarlı dokunaçlar vardır. Bu reseptörlerden gelen dürtüler, analiz için aplicia’nın beynine iletilir. Koku işleme ağı, hayvanın uygun bir yiyecekle temas kurduğunu belirlerse (yumuşakçalar yiyeceğin kokusunu almayı öğrenebilir), beslenme davranışını kontrol eden dahili nöronlara sinyaller gönderir. Ancak bu iç nöronlar aynı zamanda beynin diğer bölümlerinden beslenme karşıtı sinyaller alırlar. Örneğin, kaçma beslenmeye göre önceliklidir, böylece hayvan bir tehlike algıladığında beslenme gibi düşük öncelikli faaliyetler durdurulur. Beslemeye devam edildiğinde motor programı etkinleştirilir. Birçok omurgasız davranışında gördüğümüz gibi, merkezi sinir sistemi gangliondaki uygun devreyi engellemeyi bırakır.
Beslenme 22 kas, altı motor nöron tarafından kontrol edilir. Bu motor nöronlar, sırayla, iki antagonistik “baskın” ara nöron grubu, vericiler ve bağlayıcılar tarafından koordine edilir. İletki ve retraktör kasları, sırasıyla ekstansör ve fleksör kaslara benzer. İletkinin dahili nöronları otomatik olarak her üç veya dört saniyede bir ritmik darbeler yayar. Bu hücrelerden gelen sinyaller, belirli bir sırayla, ağzı açan ve yiyeceği ısırmak için “dişleri” aşağı indiren kasları harekete geçirir. Bu hücreler aynı zamanda bağlayıcı motor nöronları ve bunların ara nöronlarını da inhibe eder.
Derin internöronların ritmik döngüsü onları sessiz faza getirdiğinde, iletkinin internöronlarının inhibisyonunu durdururlar. İnhibisyon olmadığında, sürekli impuls yayan bu hücreler, bu noktada bağlayıcı motor nöronları aktive eder ve iletki kaslarını kontrol edenleri inhibe eder. Böylece kasların kasılma sırasını ısırmak ve ağzı kapatmak için değiştirirler.
Alpysia’nın besleme devresi, tipik bir sürüş programının üç temel özelliğini sergiler:
1- Program, farklı kasların çalışmasını koordine eden bağımsız bir devre üzerinden yürütülür.
2. Bağımsızken, duyusal geri bildirim kullanarak davranışı otomatik olarak ayarlar. Bu örnekte, uygun olduğu yerde açılış aşamasını uzatır ve güçlendirir.
3. Beynin doğrudan merkezi kontrolü altındadır. Aplysia’da beyin, kemosensitif hücrelerden ve “açlık” reseptörlerinden gelen bilgileri entegre eder ve gerektiğinde bu motor programını etkinleştirir. Aynı şekilde beyin, başka davranışlar gerektiğinde örneğin yırtıcı bir denizyıldızından kaçmak için beslenme davranışını durdurabilir. Beyin aynı zamanda davranış sıklığını da değiştirebilir; Kurs süresi ihtiyaca göre uzatılabilir veya kısaltılabilir.
Aplysia taşıyıcıları ve bastırıcıları gibi iki yönetici hücre grubu kullanma stratejisine karşılıklı engelleme denir. Bu yöntemin, görünüşte tüm ritmik davranışlar için sistem olduğu düşünülmektedir. Motor programlar ayrıca yutma ve gülme gibi ritmik olmayan davranışları da gerçekleştirir. Bir kuş evi inşa etmek veya sessizce yaklaşan bir kedi, kovalamak, yakalamak, öldürmek, dilimlemek ve bir fareyi yemek gibi daha karmaşık davranışları düzenlemeye yönelik stratejiler, birçok ayrık motor programın oluşturulmasını ve bunları koordine etmek için üst düzey devrelerin oluşturulmasını gerektirir. . Bir organizmanın davranışına, Alplysia’da bulunanlara benzer etkileşimli nöral devre gruplarının aracılık ettiğinin fark edilmesi, modern nörofizyolojinin en büyük başarısıdır.
kaynak:
https://www.sciencedirect.com
yazar: bronzlaştırıcı tonik
Diğer gönderilerimize göz at
[wpcin-random-posts]